




Choroby genetyczne bydła
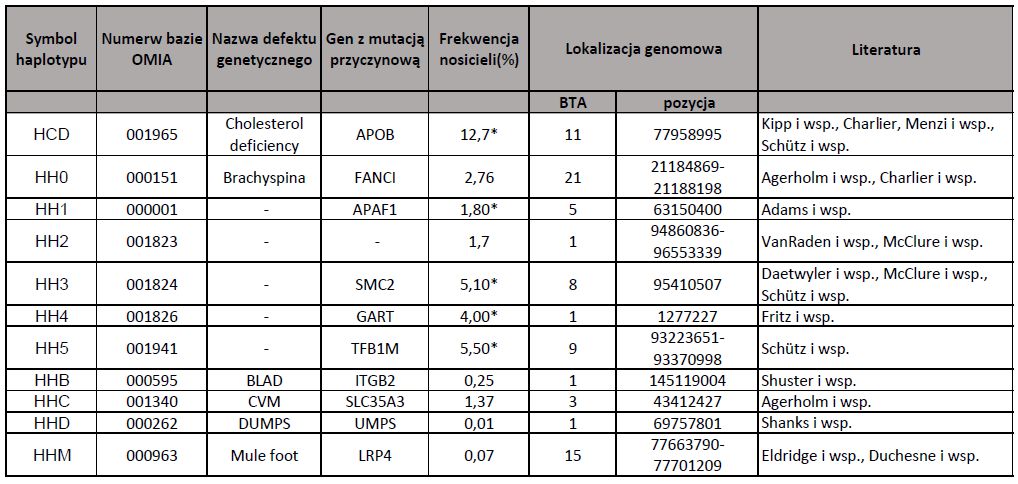
Choroby genetyczne bydła są najczęściej warunkowane przez różne haplotypy. Termin „haplotyp” odnosi się do grupy alleli, które znajdują się w pobliskich pozycjach na chromosomie i są zwykle dziedziczone razem. Nowoczesne narzędzia genomiczne zostały użyte do identyfikacji tysięcy haplotypów na każdym chromosomie. Każdy ma pozytywny, neutralny lub negatywny związek z produkcją, konformacją, zdrowiem i płodnością.
Brachyspina
Brachyspina jest warunkowana przez haplotyp HH0. Pojawiła się po raz pierwszy jako mutacja genetyczna u buhaja Sweet-Haven Tradition (o. Elevation), urodzonego w 1974 r. Jego synowie to między innymi tak znane buhaje jak Bis-May Tradition Cleitus, Rothrock Tradition Leadmon, United Nick, które widnieją w rodowodach wielu uznanych i szeroko stosowanych w hodowli buhajów. Brachyspina została opisana po raz pierwszy dopiero w 2007 r. jako pojedynczy recesywny defekt genetyczny. Oznacza to, że wada ta ujawnia się tylko wtedy, gdy zarówno buhaj jak i kojarzona z nim krowa są nosicielami zmutowanego genu i dotyczy to przeciętnie 25% potomstwa. Pozostałe 50% potomstwa to nosiciele tego defektu, kolejne 25% to zwierzęta w pełni zdrowe. Samo nosicielstwo jest bezobjawowe, dlatego do niedawna jego wykrycie przy użyciu dostępnych technik nie było możliwe. Jednak szybki postęp nauk genetycznych w zakresie selekcji genomowej i markerów genetycznych sprawił, że zidentyfikowano buhaje będące nosicielami recesywnego genu, a Światowa Federacja Bydła Holsztyńsko-Fryzyjskiego (WHFF) dla oznaczenia nosicieli tego defektu genetycznego przyjęła skrót BY. Coraz częściej i coraz więcej buhajów jest już przebadanych na nosicielstwo Brachyspiny, a skrót BY- (wolny od nosicielstwa) zaczyna być ważnym wyznacznikiem przy wyborze buhaja do dalszej hodowli.
Objawy Brachyspiny to: poronienia występujące u 0,16% krów w populacji, martwo urodzone cielęta po normalnie przebiegającej ciąży, niedorozwój narządów wewnętrznych takich jak np. nerki, serce, skrócony kręgosłup, wydłużone kończyny, bardzo niska waga urodzeniowa cieląt.
Niedobór syntetazy monofosforanu urydyny (DUMPS)
DUMPS jest warunkowany przez haplotyp HHD. Jest to letalna recesywna mutacja genu syntetazy urydynomonofosforanowej. Gen odpowiedzialny za tą mutację jest też nazywany „genem wczesnej zamieralności zarodków” gdyż homozygotyczne zarodki zamierają około 40 dnia ciąży w okresie ich implantowania się w macicy. Skutkiem tego krowa po opuszczeniu jednej rui wchodzi w kolejny cykl rujowy, co wydłuża jej okres międzywycieleniowy i ma istotny wpływ na rachunek ekonomiczny. DUMPS jest jedną z wcześniej wykrytych wad – w USA już w 1988 roku wprowadzono oficjalnie testowanie bydła w celu wykrycia nosicieli DUMPS. Za jednego z głównych nosicieli mutacji DUMPS uznaje się buhaja Skokie Sensation Ned, urodzonego w 1957 roku, z którego linii pochodzi większość znanych rozpłodników przekazujących tę wadę. W ostatnich latach największym światowym nosicielem mutacji DUMPS był amerykański buhaj Happy Herd Beautician i to za jego pośrednictwem nastąpiło rozpowszechnienie się tej anomalii na świecie. Nosicielstwo oznaczane jest w katalogach buhajów symbolem DP, a buhaje wolne od nosicielstwa oznacza się skrótem DP-.
Wrodzony niedobór leukocytarnych cząsteczek adhezyjnych lub jeszcze inaczej syndrom wrodzonego braku odporności (BLAD)
BLAD jest warunkowany przez haplotyp HHB. Jest to letalna choroba genetyczna o charakterze recesywnym, występującą u bydła HF. Jego istotą jest zanik funkcji obronnych leukocytów, tj. białych ciałek krwi. Cielęta obciążone tą mutacją w formie homozygotycznej charakteryzują się brakiem odporności i cierpią na częste, powtarzające się infekcje bakteryjne, owrzodzenia wewnątrz i wokół jamy ustnej, zapalenia przewodu pokarmowego uniemożliwiające właściwe pobieranie pokarmu, biegunki, infekcje układu oddechowego, a masa ich ciała stanowi około 60% normy. Wymienione objawy prowadzą do opóźnionego wzrostu i rozwoju cieląt, które zwykle padają w pierwszych miesiącach życia, na skutek infekcji wyniszczających organizm. Rozpowszechnianie mutacji BLAD następuje przez kojarzenie ze sobą heterozygotycznych nosicieli zmutowanego allelu, u których nie obserwuje się fenotypowych objawów upośledzenia funkcji immunologicznych. Sprzyja temu także fakt, że nosiciele mutacji charakteryzują się często wysoką wartością hodowlaną. Wykazano między innymi, że krowy nosicielki mutacji produkowały więcej mleka i białka niż ich półsiostry o prawidłowym genotypie. Zespół BLAD u bydła po raz pierwszy opisano w 1983 roku. Przyjęto, że buhaje będące nosicielami BLAD oznacza się w katalogach symbolem BL, a wolne od nosicielstwa BL-.
Zespół zniekształceń kręgosłupa (CVM - Complex Vertebral Malformation)
CVM jest warunkowany przez haplotyp HHC. Jest to letalne schorzenie o charakterze recesywnym występującym u bydła HF. Wyróżnia się dwie formy deformacji cieląt. Pierwsza z nich dotyczy układu kostnego. Ciała cieląt donoszonych, ale martwo urodzonych, a także płody poronione przed 260 dniem ciąży, charakteryzuje skrócony odcinek szyjny i piersiowy kręgosłupa, obustronne, symetryczne zesztywnienia nadgarstków, śródręcza i śródstopia, przykurcz palców w stawach pęcinowych, brak niektórych kręgów oraz wadliwe połączenia żeber z kręgami. Deformacje występujące w obrębie kręgosłupa, powodują uszkodzenia lub urazy rdzenia kręgowego, a nasilenie zmian patologicznych bywa różne u różnych osobników. Druga forma anomalii dotyczy układu krwionośnego i oddechowego. Negatywne skutki defektu CVM mogą występować w każdym okresie rozwoju prenatalnego, objawiając się resorpcją zarodków lub poronieniami przed 260 dniem ciąży, a także wydalaniem martwych płodów przeważnie 1–2 tygodnie przed planowaną datą wycielenia. Pierwszy przypadek urodzenia martwego cielęcia z objawami CVM odnotowano w Danii w 2000 roku, kolejne martwo urodzone cielęta z tym schorzeniem zarejestrowano w USA, Holandii i Wielkiej Brytanii. Ze względu na fakt, że CVM przynosi znaczne straty w reprodukcji bydła HF, w wielu krajach wprowadzono programy wykrywania nosicieli tego defektu wśród buhajów wykorzystywanych w sztucznej inseminacji przy pomocy testu DNA. Buhaje będące nosicielami mutacji oznacza się symbolem CV umieszczonym przy nazwie buhaja, natomiast osobnika wolnego od CVM oznacza się skrótem CV-.
Defekt genetyczny zwany "Deficytem cholesterolu CD"
Deficyt cholesterolu CD jest warunkowany przez haplotyp HCD. W stadach bydła często zdarza się, że cielęta padają ze względu na niedostateczny rozwój, utratę apetytu i wyniszczające biegunki. Pomimo starań weterynaryjnych stan tych cieląt nie poprawia się. Objawy choroby to przede wszystkim przewlekła, nieuleczalna biegunka, niedowaga, częste powikłania to zapalenie płuc i ich obrzęk. Test na patogeny wychodzi negatywny, co dowodzi, że musi być inna przyczyna. Zaobserwowano również, że bliźniaki odchowywane w różnych gospodarstwach wykazują te same objawy chorobowe.
Naukowcy przeprowadzili badania patologiczne na cielętach, które już w chwili urodzenia były słabsze. Ich wiek w dniu upadku wynosił od 1 do 5 miesięcy, a waga 36,5 - 59 kg, wszystkie były normalnej wielkości ciała. Nie znaleziono bezpośredniej przyczyny śmierci, najprawdopodobniej zwierzęta padły z głodu. Wynik badania patologicznego wykazał, że wszystkie zapasy tkanki tłuszczowej były wyczerpane, nie było nawet tłuszczu w rdzeniu kręgowym, co może sugerować kłopoty z jego prawidłowym wchłanianiem. Ostatnio grupa badawcza składająca się z przedstawicieli Vit z Verden, Instytutu Weterynarii w Kiel, Szkoły Wyższej w Hanowerze oraz Uniwersytetu Technicznego w Monachium odkryła genetyczną przyczynę tych upadków. Grupa badawcza zebrała dużo informacji o cielętach o podobnych rodowodach, które cierpią na niepoddającą się terapii i prowadzącą do śmierci biegunkę oraz charakteryzują się niedostatecznym rozwojem. Dzięki udanej współpracy środowisk weterynaryjnych (MASTERRIND) odkryto genetyczną przyczynę tej nieuleczalnej choroby. W oparciu o współczesne możliwości analizy genomu oraz rozszerzoną wiedzę o sposobach dziedziczenia u bydła odkryto w genomie haplotyp bezpośrednio powiązany z tą chorobą. Ponadto, grupa odkryła związek pomiędzy zmienionym odcinkiem genomu, a zaburzonym mechanizmem wchłaniania cholesterolu, a ten nowy defekt genetyczny nazwano „Deficytem cholesterolu (CD)”.
Badacze przeprowadzili analizę rodowodów, która wykazała, że wszystkie cielęta, które cierpiały i padły na CD w swoim rodowodzie posiadały (zarówno od strony ojcowskiej, jak i matczynej) urodzonego w 1991 roku kanadyjskiego buhaja Maughlin Storm. Ponadto wszystkie te cielęta były homozygotyczne dla konkretnego haplotypu (nazwanego CD) na chromosomie 11. Jednak po przeprowadzeniu pełnej analizy populacji na haplotyp CD okazało się, że było również sporo homozygotycznych zwierząt całkiem zdrowych, które pochodziły od jednego przodka (którego Storm ma również w rodowodzie). Chodzi tutaj o urodzonego w 1966 roku Fairlea Royal Mark. Jest więc oczywiste, że w populacji znajdują się dwa warianty haplotypu CD: wadliwy wariant po buhaju Storm i niegroźny wariant po buhaju i F.R.Mark. Mutacja musiała więc nastąpić u buhaja Storm lub któregoś z buhajów między generacją Storm, a Fairlea Royal Mark. Dobrą metodą oszacowania jaki wariant haplotypu CD posiada buhaj–nosiciel jest analiza rodowodu. Jeśli buhaj nie ma w rodowodzie buhaja Storm posiada on wariant niegroźny. Jeśli jego rodowód zawiera tylko buhaja Storm, a nie ma F.R.Mark to musi być nosicielem zmutowanego haplotypu. Jeśli zawiera oba te buhaje może posiadać oba warianty.
Fakt, że "podział" niegroźnego i wywołującego chorobę haplotypu miał miejsce przed wieloma pokoleniami spowodował, że oba te warianty są rozpowszechnione w populacji i ich nosiciele posiadają w rodowodach zarówno buhaja Storm, jak i F.R.Mark. Przykładem może być znana kanadyjska matka buhajów Comestar Laurie Sheik (prawnuczka buhaja F.R.Mark), która jest nosicielką oczywiście niegroźnego wariantu haplotypu, a ze względu na jej szerokie użycie w hodowli haplotyp ten został szeroko rozpowszechniony w populacji HF.
Obecnie jest możliwość przeprowadzenia testu, który z prawdopodobieństwem 98% pozwala wykryć haplotyp CD w genomie. Problem jest z testem, który pozwala określić wariant tego haplotypu u zwierzęcia-nosiciela, ponieważ takie zwierzę może posiadać wariant groźny lub nieszkodliwy. Obecnie pracuje się na tym, aby opracować test, który będzie w stanie rozróżnić oba te warianty.
W Niemczech rodzi się 1 800 000 cieląt rasy HF. Z tego 3400 sztuk jest homozygotyczna pod względem haplotypu CD, co powoduje, że chorują i padają. Nosicielami haplotypu CD jest 8.7% populacji HF czyli ok. 157000 sztuk. To powoduje straty w wysokości 1,3 mln €, czyli w przeliczeniu na jedno cielę 400 € (średni czas życia i utrzymania cielęcia to 85 dni, do tego koszty weterynaryjne). Wszystkie martwe zwierzęta homozygotyczne pod względem tego haplotypu po obu stronach rodowodu mają buhaja Maughlin Storm.
Buhaje będące nosicielami mutacji oznacza się symbolem CD+ umieszczonym przy nazwie buhaja, natomiast osobnika wolnego od CD oznacza się skrótem CD-.
Haplotypy obniżające płodność bydła HF
Ostatnio odkryto wiele haplotypów, które negatywnie wpływają na płodność bydła holsztyńsko-fryzyjskiego. Znane są jako HH1, HH2, HH3, HH4, HH5 i HH6. Uznanie haplotypu jako istotnego dla cech płodności następuje po spełnieniu następujących warunków: osobniki homozygotyczne rodzą się martwe lub występuje wyraźna redukcja skuteczności zapłodnienia, lub wzrost liczby poronień wskutek kojarzenia buhaja nosiciela z krową pochodzącą po buhaju nosicielu. Uznaje się, że HH dziedziczą się w sposób autosomalny recesywny. Efekty fenotypowe i ekonomiczne jakie wywołują haplotypy w stanie homozygotycznym są zróżnicowane i zależą głównie od ich frekwencji oraz fazy ciąży, w której ujawnia się ich działanie. Im wyższa częstość występowania haplotypu i im późniejsza faza ciąży, tym starty wywołane haplotypami są większe. Objawy kliniczne mają miejsce zazwyczaj przed 60-100 dniem ciąży (HH2, HH3, HH4, HH5 i HH6) lub we wszystkich stadiach ciąży (HH1).
Haplotypy HH1 – HH4 to szereg haplotypów, które upośledzają płodność w populacji holsztyno-fryzów. Są odpowiedzialne za zaburzenia reprodukcyjne. Haplotypy te w stanie homozygotycznym skutkują nieudanym zapłodnieniem lub wczesną śmiercią embrionalną. Używanie nasienia od byków-nosicieli tych haplotypów zazwyczaj powoduje około 3% niższy wskaźnik zapłodnienia.
Haplotyp HH5 jest sprawcą mutacji w postaci rozległej delecji wielkości 138 tysięcy par zasad, która powoduje usunięcie genu TFB1M kodującego enzym transferazę 1-dimetyloadenozynową. Brak tego enzymu destabilizuje rybosomy i prowadzi do zaniku translacji białek, a w konsekwencji do zamierania komórek, co prowadzi do śmierci płodu.
Haplotyp HH6 został wykryty stosunkowo niedawno, a w marcu 2018 roku w czasopiśmie „Journal of Dairy Science” ukazała się praca Sebastiana Fritz i współpracowników opisująca molekularną przyczynę tego defektu genetycznego. Zespół ten przebadał 250 tys. osobników rasy holsztyńskiej i zidentyfikował nowy haplotyp HH6, wykazując jego istotny negatywny wpływ na wskaźnik zapłodnienia (Conception Rate) oraz niepowtarzalność rui w 56. dniu. Badaczom udało się zmapować region chromosomu 16 wielkości 1,1 mln par nukleotydów i przeanalizować ten obszar, porównując sekwencje 12 nosicieli i 284 buhajów zdrowych. W ostatecznym efekcie powoduje to całkowitą dysfunkcję białka SDE2 i ‒ co za tym idzie ‒ poważne konsekwencje dla rozwoju zarodka. Przypuszcza się, że dysfunkcja białka SDE2 prowadzi do zamierania zarodków po kilku podziałach mitotycznych, co koresponduje z zanikiem żywych homozygot recesywnych oraz podwyższonym wskaźnikiem niepowtarzalności rui po 56. dniu. Badania populacyjne u ponad 46 tys. zwierząt wykluczyły występowanie mutacji u innych ras niż holsztyńska. Badania wykonane na populacji 29 tys. zwierząt rasy holsztyńsko-fryzyjskiej wykazały brak osobników w stanie homozygotycznym, co upoważnia do stwierdzenia, że omawiana mutacja ma charakter letalny. Kojarzenie dwóch nosicieli HH6 prowadzi do spadku wskaźnika zapłodnienia o 7-11%.